生体膜脂質の機能を探る
はじめに
全ての生物は細胞からなり,個々の細胞は細胞膜という生体膜によって仕切られ,細胞内に存在する細胞小器官も生体膜によって区画化されている.これらの生体膜は細胞が生きていくために必要な構造体である.これらの生体膜のおもな構成成分は脂質で,脂質が形成する脂質二重層が生体膜の基本構造となっている.細胞が示すほとんどの生命現象は生体膜に依存していることから,それらの生命現象を分子レベルで解明するには脂質の働きを理解することが必要である.生体膜を構成する脂質は極性脂質と呼ばれており,極性脂質にはグリセロールを骨格とするグリセロ脂質やスフィンゴイド塩基を骨格とするスフィンゴ脂質があり,また各々の極性脂質には糖を含む糖脂質やリンを含むリン脂質など,数十種類の脂質クラスが知られている.さらに,各クラスの脂質分子には脂肪酸が結合しており,その脂肪酸には飽和脂肪酸や不飽和脂肪酸,また水酸基などによって修飾されている脂肪酸も存在し,不飽和脂肪酸には二重結合の数や位置が違うものが多数知られている.したがって,細胞には構造の異なる多様な脂質分子が存在していることになり,その分子の種類は数千にものぼる.なぜ,そのような多様な脂質分子が必要なのであろうか?脂質がただ単に細胞や細胞小器官の内と外を区画化する仕切りとしてのみ働いているのであれば,これだけの種類の脂質が存在する必要はないであろう.脂質の種類や組成は膜ごとに異なっており,脂質が各々の生体膜のもつ機能を発揮する上で何らかの重要な働きをもつことが容易に想像できる.筆者らはこの多様な脂質分子の機能について,シアノバクテリアや高等植物といった光合成生物を用いて解析している.
光合成における機能
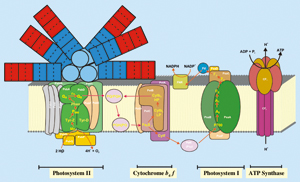 図1 チラコイド膜でおこる光合成の初期過程
図1 チラコイド膜でおこる光合成の初期過程光合成は,植物が光エネルギーを利用して水を酸化するとともに空気中の炭酸ガスを固定して糖やでんぷんを合成する反応であり,地球上のほとんどの生物の生存に必要なエネルギーの源はこの光合成によって賄われている.この光合成の初期過程の場は,シアノバクテリアや植物の葉緑体に存在するチラコイド膜であり,この膜には光エネルギーを化学エネルギーに変換する光化学系II,チトクロムb6/f,光化学系I,ATP合成酵素と呼ばれる超分子複合体(光合成装置)が存在する(図1).このチラコイド膜は光合成の場であり,しかも細胞に存在する膜の大部分を占めるため,質的,量的に最も注目されるべき生体膜である.チラコイド膜は他の生体膜とは異なり,糖脂質が主成分であるという特徴をもっている(図2).この膜が糖脂質を主成分とする理由は,光合成生物にとって大変貴重なリンを節約するためであり,リン脂質の糖脂質への変換が進化の過程でおこったと考えられる.しかし,チラコイド膜にはホスファチジルグリセロール(PG)という脂質が唯一のリン脂質として存在している.
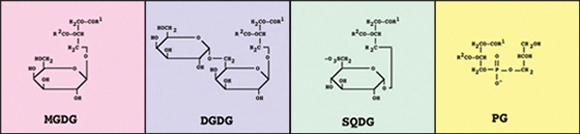 図2 チラコイド膜に存在する脂質の構造
図2 チラコイド膜に存在する脂質の構造筆者らは,このPGという脂質クラスに注目している.この脂質が糖脂質では担うことのできない特別な機能をもつのではないかと予想したからである.シアノバクテリアSynechocystis sp. PCC6803(図3)からPG合成欠損株(PGの合成に関わっている遺伝子を破壊した変異株)を作製し,その変異株を用いてPGの機能を解析している.この株はPGを全く合成できず,通常の培地では増殖できない.しかし,PGを培地に加えておくと,それを細胞内に取り込んで増殖することが可能である(図4).このため,この株はPGの機能を解析するのに大変都合のよい実験材料である.PGを加えた培地で培養した変異株の細胞をPGが含まれていない培地に移すと,細胞分裂にともなってPGの含有量が低下し,数回分裂した後に細胞の増殖が停止する.このとき,PGの含有量の低下に伴って光合成活性も低下し,PGを再添加すると活性が速やかに回復する.このことは,PGが増殖および光合成に必要であることを示している.PGが光合成になぜ必要なのかについては,詳細な解析の結果,PGが光化学系IIにおけるQAからQBへの電子伝達を維持するのに必要であることが明らかとなっている.また,この変異株はPGを含まない培地に移すと,初期段階では光化学系IIの活性のみが低下するが,さらに長く培養すると,光化学系Iの活性も低下する.このとき,両活性の低下とともに,光化学系II複合体および光化学系I複合体のモノマーが蓄積することから,PG が光合成活性の維持に必要で,複合体のオリゴマー化や安定化に寄与していることがわかってきている.
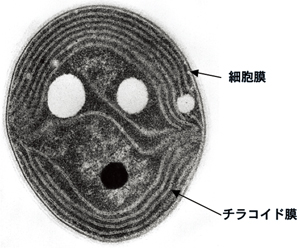 図3 シアノバクテリアSynechocystis sp. PCC6803の電子顕微鏡写真
図3 シアノバクテリアSynechocystis sp. PCC6803の電子顕微鏡写真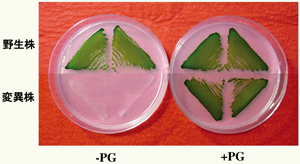 図4 寒天培地上で増殖させたシアノバクテリアSynechocystis sp. PCC6803の野生株とPG合成欠損株
図4 寒天培地上で増殖させたシアノバクテリアSynechocystis sp. PCC6803の野生株とPG合成欠損株タンパク質の脂質修飾における機能
PGは光合成において重要な働きを担っているが,PGはシアノバクテリアのリポタンパク質と呼ばれる一群のタンパク質の脂質修飾にも関わっている.シアノバクテリアを含む全てのバクテリアには,リポタンパク質と呼ばれる脂質によって修飾されるタンパク質が存在する.それらのタンパク質は前駆体として合成され,脂質修飾などのプロセシングを受けて成熟型のタンパク質になる.リポタンパク質は脂質で修飾されることにより,膜に結合できるようになる.前駆体は,まず,N-末端領域にあるシステイン残基のSH基でPGによる修飾(ジアシルグリセロール化)を受けてからシグナルペプチダーゼIIによってシグナルペプチド部分が切断され,さらにN末端のシステイン残基のアミノ基がアシル基によって修飾されて成熟型となる.リポタンパク質の機能は,ペプチドグリカンの構造維持,物質輸送,シグナル伝達など,多岐にわたっている.シアノバクテリアSynechocystis sp. PCC6803のゲノムの塩基配列からリポタンパク質の遺伝子を検索すると,このシアノバクテリアには約40の遺伝子が存在することがわかっている.このことから,シアノバクテリアには多くのリポタンパク質が存在し,PGを介した脂質修飾がそれらのリポタンパク質の機能を発揮する上で重要な働きをもつことが考えられる.各々のリポタンパク質がなぜ脂質修飾されなければいけないのか,その理由を解析するために脂質修飾できない変異株の作製を進めている.それらの変異株を調べることにより,リポタンパク質がPGを用いて脂質修飾されることの意義がわかるものと期待される.
葉緑体の分化における機能
高等植物であるシロイヌナズナについてもシアノバクテリアと同様の解析を行っている.シロイヌナズナから葉緑体でのPGの合成に関わっている遺伝子(PGP1)が破壊された2つの変異株の分離を行なった.これらの変異株では,PGP1遺伝子が破壊されることにより,葉緑体でのPG合成が欠損している.分離した2つの変異株はアルビノに近いpale greenの葉を形成し,土の上では生育できず,ショ糖を加えた寒天培地でのみ生育することができる(図5).また,変異株は正常な形の葉を形成することができず,細胞内には葉緑体が見出されない.これらの観察結果は,PGが植物の光独立栄養成長に必須であり,葉緑体の分化や葉の形成においても重要な働きを担っていることを示している.現在,この変異株の性質をさらに詳しく調べており,それらの解析から葉緑体の分化や葉の形成におけるPGの機能がわかってくるものと考えている.
 図5 寒天培地上で生育させたシロイヌナズナの野生株とPG合成欠損株
図5 寒天培地上で生育させたシロイヌナズナの野生株とPG合成欠損株おわりに
このように,PGはシアノバクテリアや高等植物において大変重要な機能をもつことがわかってきた.生物には,PG以外にも多種多様な脂質分子が存在し,それらの機能を解析することは大変重要である.その解析には,筆者らがPGについて行なっているように,変異株を用いることが大変有効である.最近,多くの生物のゲノムの塩基配列が決定され,脂質の合成に関わっている遺伝子のほとんどが同定されている.それらの遺伝子の発現を人為的に操作することで,特定の脂質分子の合成を制御することが可能になってきている.今後,そのような遺伝子操作によって作製された変異株を使った解析によって,新たな脂質の機能が次々と明らかになってくるものと期待される.